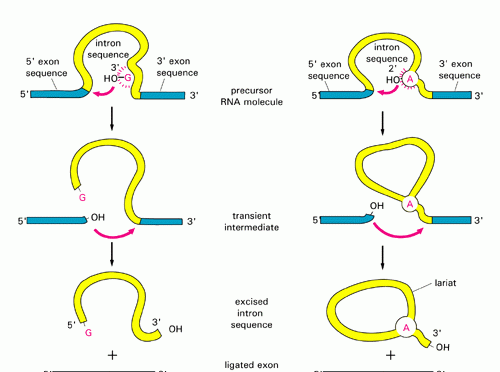
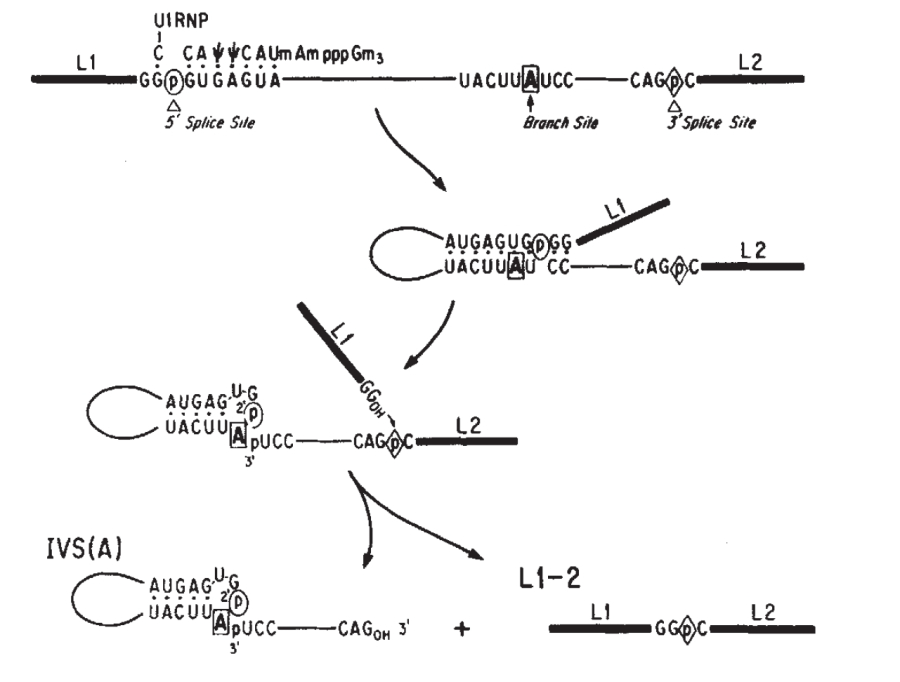
SPLICING
Fibre Rope Termination
For a rope to transmit force it needs a termination, whether it is a permanent attachment, such as a splice, socket or mechanical grip, or a temporary fix, such as a knot or wraps around a post.
An effective termination is essential to almost every application that puts a rope under tension.
Splicing
Splicing can be used in three ways. An eye spice puts a loop in the end of a rope. An end-to-end splice can either join two ropes together or, finally, join two ends of the same rope to make a circular grommet.
Fibre rope splicing is a skill that must be learned. The simpler splices can be produced by carefully following a manual.
Small practices that are learned from experience or by testing of the splices are often needed to produce the best results.
A well-practiced expert can make splices that match rope strength.
The most common and one of the most dependable methods of terminations of the fibre ropes is an eye splice, which can be placed round a suitable fitting.
Splices are made by separating the strands at the ends of the rope from the structure, bending the rope into a loop and tucking the separated strands into the body of the rope.
Another approach is to separate the strands, create an eye, and then braid them over the exterior of the rope; this over-braid will tighten and grip as tension is applied at the eye.
An intermediate stage of tucking the strands is an eight-strand plaited rope. There is enough grip on the tucked strands to hold considerable tension, usually to the maximum breaking strength of the rope.